Вирус гепатита с: кач. рнк обнаружено
Содержание:
Разница между ДНК и РНК-вирусами
Определение
ДНК-вирусы: ДНК-вирусы относятся к вирусам, генетическая информация которых хранится в форме ДНК.
РНК Вирусы: РНК-вирусы относятся к вирусам, генетическая информация которых хранится в форме РНК.
геном
ДНК-вирусы: ДНК-вирусы содержат ДНК в качестве своего генетического материала.
Вирусы РНК: РНК-вирусы содержат РНК в качестве своего генетического материала.
Дважды многожильный / Однонитевый
ДНК-вирусы: Двухцепочечные ДНК-вирусы встречаются чаще, чем одноцепочечные ДНК-вирусы.
Вирусы РНК: Однонитевые РНК-вирусы встречаются чаще, чем двухцепочечные РНК-вирусы.
копирование
ДНК-вирусы: Вирусная ДНК реплицируется внутри ядра клетки-хозяина.
Вирусы РНК: Вирусная РНК сначала транскрибируется, а затем реплицируется в цитоплазме.
Синтез белка
ДНК-вирусы: Вирусная ДНК сначала транскрибируется в РНК, а затем мРНК транслируется в вирусные белки.
РНК Вирусы: РНК-вирусы могут обходить транскрипцию во время синтеза белка, поскольку они уже содержат РНК в геноме.
стабильность
ДНК-вирусы: ДНК-вирусы стабильны из-за более низкой частоты мутаций.
РНК Вирусы: РНК-вирусы нестабильны из-за более высокой частоты мутаций.
Верность репликации
ДНК-вирусы: ДНК-вирусы показывают точную репликацию.
РНК Вирусы: РНК-вирусы показывают подверженную ошибкам репликацию.
ДНК-вирусы: ДНК-вирусы содержат большой геном.
Вирусы РНК: РНК-вирусы содержат небольшой геном.
прокапсида
ДНК-вирусы: Вновь синтезированная вирусная ДНК упаковывается в предварительно образованный капсид, называемый прокапсид.
Вирусы РНК: Вновь синтезированная вирусная РНК должна быть защищена от деградации, поскольку у РНК-вирусов не образуется прокапсид.
Типы
ДНК-вирусы: Классы I, II и VII Балтиморской классификации вирусов являются ДНК-вирусами.
Вирусы РНК: Классы III, IV, V и VI Балтиморской классификации вирусов являются РНК-вирусами.
Примеры
ДНК-вирусы: Аденовирусы, герпесвирусы, поксвирусы, парвовирусы и гепаднавирусы являются примерами ДНК-вирусов.
Вирусы РНК: Реовирусы, пикорнавирусы, тогавирусы, ортомиксовирусы, рабдовирусы и ретровирусы являются примерами РНК-вирусов.
Сопутствующие заболевания
ДНК-вирусы: Оспа, герпес и ветряная оспа являются заболеваниями ДНК-вирусов.
Вирусы РНК: СПИД, геморрагическая лихорадка Эбола, ОРВИ, простуда, грипп, гепатит С, лихорадка Западного Нила, полиомиелит и корь — вот некоторые заболевания, вызываемые вирусами РНК.
Заключение
ДНК и РНК-вирусы — это два типа вирусов с различным типом генетического материала. ДНК-вирусы содержат ДНК в геноме, а РНК-вирусы содержат РНК. Репликация ДНК-вирусов происходит внутри ядра хозяина, в то время как происходит в цитоплазме хозяина у РНК-вирусов. Основное различие между ДНК и РНК-вирусами заключается в типе генетического материала и репликации внутри ядра.
Как убедиться в достоверности обследований?
И ложноположительный результат, и положительный, но без наличия каких-либо симптомов должны подтолкнуть пациента получить достоверные сведения о состоянии своего здоровья. Для этого нужно делать другой анализ – «ПЦР качественный» или «ПРЦ гепатита C». Этот анализ выявляет не антитела, а непосредственно РНК вируса – т.е. наличие его активной формы в организме пациента в данный момент времени.
Если делать анализ на антитела правильно и он показал положительный результат, а ПЦР качественный – отрицательный, это значит, что течение болезни перешло в скрытую форму, или она излечилась сама собой.
В этом случае делать другие исследования не стоит, и лечение пациенту не нужно, но необходимо повторять анализ ПЦР по крайней мере раз в год, для того чтобы выявить переход вируса в активную форму и начало болезни. Также желательно перестать принимать алкоголь и жирную пищу, чтобы исключить все факторы риска поражения печени.
В этом случае пациент просто не замечает ни начала, ни окончания течения заболевания – возможны лишь общие признаки недомогания, которые можно списать на стресс или простуду. Тем не менее, если у пациента обнаружили антитела к гепатиту, он должен в течение всей жизни ежегодно сдавать анализы, чтобы убедиться в отсутствии перехода вируса в активную форму.
Переход вируса в хроническую форму тоже не представляет опасности для самого пациента – он может так же, как и здоровый человек прожить долгую плодотворную жизнь, не страдая ни от каких проявлений. Разумеется, это возможно только при соблюдении рекомендаций врача и регулярной диагностике методом ПЦР гепатита C.
Человеку с положительным результатом ПЦР на гепатит C стоит помнить о мерах предосторожности в общении с окружающими. Вирус гепатита C передается через кровь и при попадании частицы крови в другие жидкости тела, например, в слюну, если во рту есть незначительная ранка
Значит, для того чтобы не заразить вирусом своих близких, следует сказать им «у меня гепатит С» и придерживаться таких мер предосторожности:
Вирус гепатита C передается через кровь и при попадании частицы крови в другие жидкости тела, например, в слюну, если во рту есть незначительная ранка
Значит, для того чтобы не заразить вирусом своих близких, следует сказать им «у меня гепатит С» и придерживаться таких мер предосторожности:
- не пользоваться общими иглами (при нанесении татуировки, пирсинге, уколах);
- при порезах кухонным ножом его лезвие следует продезинфицировать;
- при ранениях кровь следует удалять с поверхностей и предметов при помощи раствора хлора, вещи стирать при высокой температуре;
- если во рту есть ранки или кровоточат десны, следует воздержаться от поцелуев;
- следует использовать барьерные методы контрацепции во время полового акта, чтобы исключить попадание крови на слизистые (во время менструации, при наличии микротрещин).
Гепатит С не передается:
- воздушно-капельным путем;
- при рукопожатиях, объятиях;
- при использовании общих вещей и посуды при условии соблюдения правил, указанных выше.
Способы лечения
Единственным действенным способом вылечить вирус гепатита С, как правило, оказывается сочетание 2-х медпрепаратов:
- интерферона-альфа совместно с рибавирином;
- софосбувир вместе с велпатасвиром;
- софосбувир совместно с даклатасвиром.
По отдельности эти лекарства не так эффективны. Рекомендуемые дозировки лекарств и сроки применения должны назначаться только врачом и индивидуально каждому пациенту. Лечение этими лекарствами может растянуться на срок от 6 до 12 месяцев по первой схеме и от 3 до 6 месяцев по второй и третьей схеме.
https://youtube.com/watch?v=X6UURHW60b0
Сегодня не изобретено препаратов, гарантирующих стопроцентное выздоровление от вируса. Однако при верно подобранном лечении исцеление пациентов может достигать до 98% от числа заболевших.
Значение РНК и ДНК
Когда было открыто, что такое ДНК, ее роль не была такой очевидной. Даже сегодня, несмотря на то, что раскрыто намного больше информации, остаются без ответов некоторые вопросы. А какие-то, возможно, еще даже не сформулированы.
Общеизвестное биологическое значение ДНК и РНК заключаются в том, что ДНК передает наследственную информацию, а РНК участвует в синтезе белка и кодирует белковую структуру.
Однако существуют версии, что эта молекула связана с нашей духовной жизнью. Что такое ДНК человека в этом смысле? Она содержит всю информацию о нем, его жизнедеятельности и наследственности. Метафизики считают, что опыт прошлых жизней, восстановительные функции ДНК и даже энергия Высшего «Я» — Творца, Бога содержится в ней.

По их мнению, цепочки содержат коды, касающиеся всех аспектов жизни, включая и духовную часть. Но некоторая информация, например, о восстановлении своего тела, расположена в структуре кристалла многомерного пространства, находящегося вокруг ДНК. Она представляет собой двенадцатигранник и является памятью всей жизненной силы.
Ввиду того, что человек не обременяет себя духовными знаниями, обмен информации в ДНК с кристаллической оболочкой происходит очень медленно. У среднестатистического человека он составляет всего пятнадцать процентов.

Предполагается, что это было сделано специально для сокращения жизни человека и падения на уровень дуальности. Таким образом, у человека растет кармический долг, а на планете поддерживается необходимый для некоторых сущностей уровень вибрации.
Действие РНК-полимеразы
Связывание и инициирование транскрипции
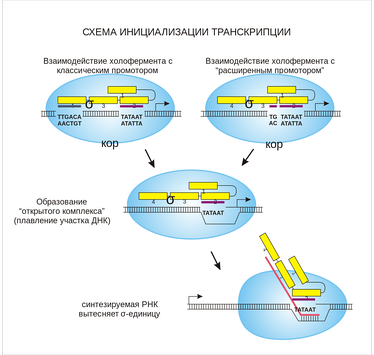 Схема инициализации транскрипции
Схема инициализации транскрипции
В связывании РНК-полимеразы участвует α-субъединица, распознающая элемент ДНК, предшествующий гену (-40…-70 шагов), и σ-фактор, распознающий участок −10…-35. Существует большое количество σ-факторов, контролирующих экспрессию генов. Например: σ70, который синтезируется в нормальных условиях и позволяет РНК-полимеразе связываться с генами, отвечающими за метаболические процессы клетки; или σ32, блокирующий связывание РНК-полимеразы с генами белков теплового шока.
После связывания с ДНК структура РНК-полимеразы превращается из закрытой в открытую. Это превращение включает в себя разделение моноспиралей ДНК с образованием раскрученного участка длиной около 13 шагов. Рибонуклеотиды затем собираются в цепочку в соответствии с базовой нитью ДНК, используемой в качестве шаблона. Суперскрученность молекул ДНК играет существенную роль в деятельности РНК-полимеразы: поскольку участок ДНК перед РНК-полимеразой раскручен, в нем существуют положительные компенсационные супервитки. Участки ДНК позади РНК-полимеразы снова закручиваются и в них присутствуют отрицательные супервитки.
Элонгация
Во время элонгационной фазы транскрипции происходит добавление рибонуклеотидов к цепи и переход от структуры РНК-полимеразного комплекса от открытой к транскрипционной. По мере сборки молекулы РНК участок ДНК перед РНК-полимеразой раскручивается далее, и 13-парный открытый комплекс превращается в 17-парный транскрипционный комплекс. В этот момент промотор (участок ДНК −10…-35 шагов) завершается, и σ-фактор отделяется от РНК-полимеразы. Это позволяет остальному РНК-полимеразному комплексу начать движение вперед, так как σ-фактор удерживал его на месте.
17-парный транскрипционный комплекс содержит гибрид ДНК и РНК, содержащий 8 пар оснований — 8-шаговый участок РНК, соединенный с шаблонной цепью ДНК. По мере выполнения транскрипции рибонуклеотиды добавляются к 3′-концу собираемой РНК, и РНК-полимеразный комплекс движется по цепи ДНК. Хотя в РНК-полимеразе не обнаружено свойств, характерных для 3′-экзонуклеазы, аналогичных проверочной деятельности ДНК-полимеразы, есть свидетельства того, что РНК-полимераза останавливается и корректирует ошибки в случаях ошибочного формирования пар оснований ДНК-РНК.
Добавление рибонуклеотидов к РНК обладает механизмом, очень близким к полимеризации ДНК. Считается, что ДНК- и РНК-полимеразы могут быть эволюционно связаны. Аспарагиновые остатки в РНК-полимеразе связываются с ионами Mg2+, которые, в свою очередь, осуществляют выравнивание фосфатных групп рибонуклеотидов: первый Mg2+ удерживает α-фосфат нуклеотидтрифосфата, подлежащего добавлению в цепочку. Это позволяет осуществить связывание нуклеотида с 3′ OH-группой конца собираемой цепочки и таким образом добавить НТФ в цепочку. Второй Mg2+ удерживает пирофосфат НТФ. Общее уравнение реакции таким образом имеет вид:
(НМФ)n + НТФ —> (НМФ)n+1 + ПФi
Терминация
Терминация транскрипции РНК может быть ρ-независимой либо ρ-зависимой.
ρ-независимая терминация осуществляется без помощи ρ-фактора. Транскрипция палиндромного участка ДНК приводит к формированию шпильки из РНК, зацикленной и связанной с самой собой. Эта шпилька богата гуанином и цитозином, что делает её более стабильной, нежели гибрид ДНК-РНК. В результате 8-парный гибрид ДНК-РНК в транскрипционном комплексе сокращается до 4-парного. В случае если эти 4 последние пары оснований составлены слабыми аденином и уридином, молекула РНК отделяется.
8.Типы РНК
В зависимости от строения и выполняемой функции различают три основных типа РНК: рибосомную, транспортную и информационную (матричную).

1. Информационная РНК
Как показали исследования, информационная РНК составляет 3-5 % от общего содержания РНК в клетке. Это одноцепочная молекула, которая образовывается в процессе транскрипции на одной из цепей молекулы ДНК. Это связано с тем, что ДНК у ядерных организмов находятся в ядре, а синтез белка происходит на рибосомах в цитоплазме, поэтому возникла необходимость в «посреднике». Функцию «посредника» выполняет матричная РНК, она передает информацию о структуре белка из ядра клеток, где находится ДНК, к рибосомам, где эта информация реализуется (см. Рис. 5).
Рис. 5. Матричная РНК (мРНК)
В зависимости от объема копируемой информации, молекула матричной РНК может иметь различную длину.
Большинство матричных РНК существуют в клетке непродолжительное время. В бактериальных клетках существование таких РНК определяется минутами, а в клетках млекопитающих (в эритроцитах) синтез гемоглобина (белка) продолжается после утраты эритроцитами ядра в течение нескольких дней.
2. Рибосомная РНК
Рибосомные РНК (см. Рис. 6) составляют 80 % от всех рибосом, присутствующих в клетке. Эти РНК синтезируются в ядрышке, а в клетке они находятся в цитоплазме, где вместе с белками образуют рибосомы. На рибосомах происходит синтез белка. Здесь «код», заключенный в матричную РНК, транслируется в аминокислотную последовательность молекулы белка.
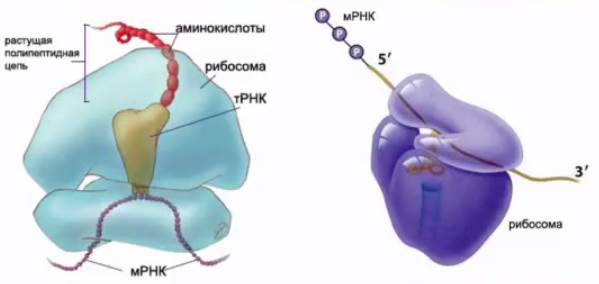
Рис. 6. Рибосомная РНК (рРНК)
3. Транспортная РНК
Транспортные РНК (см. Рис. 7) образуются в ядре на ДНК, а затем переходят в цитоплазму.
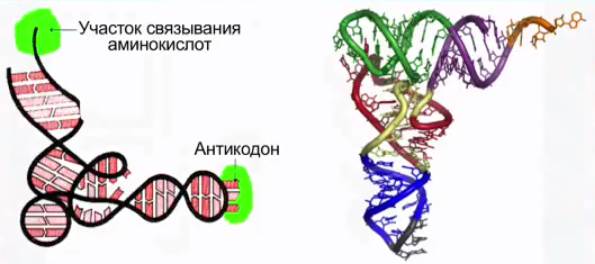
Рис. 7. Транспортная РНК (тРНК)
На долю таких РНК приходится около 10 % от общего содержания РНК в клетке. Они имеют самые короткие молекулы из 80-100 нуклеотидов.
Транспортные РНК присоединяют к себе аминокислоту и транспортируют ее к месту синтеза белка, к рибосомам.
Все известные транспортные РНК за счет комплементарного взаимодействия между азотистыми основаниями образовывают вторичную структуру, по форме напоминающую лист клевера (см. Рис. 8). В молекуле тРНК есть два активных участка – триплет антикодон на одном конце и акцепторный участок, присоединяющий аминокислоту, на другом.
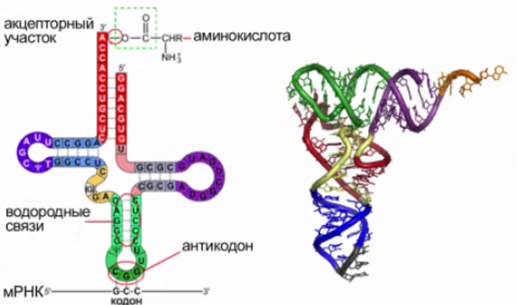
Рис. 8. Строение тРНК («клеверный лист»)
Каждой аминокислоте соответствует комбинация из трех нуклеотидов, которая носит название триплет.
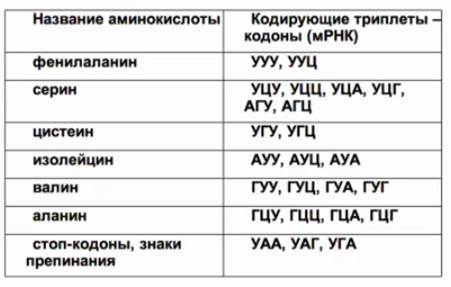
Рис. 9. Таблица генетического кода
Кодирующие аминокислоты триплеты – кодоны ДНК (см. Рис. 9) – передаются в виде информации триплетов (кодонов) мРНК. У верхушки клеверного листа тРНК располагается триплет нуклеотидов, который комплементарен соответствующему кодону мРНК (см. Рис. 10). Этот триплет различен для тРНК, переносящих разные аминокислоты, и кодирует именно ту аминокислоту, которая переносятся данной тРНК. Он получил название антикодон.
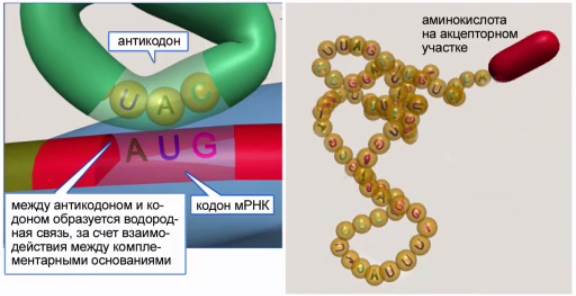
Рис. 10. тРНК
Акцепторный конец является «посадочной площадкой» для определенной аминокислоты.
Таким образом, различные типы РНК представляют собой единую функциональную систему, направленную на реализацию наследственной информации через синтез белка.
Примечания
- Nierhaus KH, Wilson DN. Protein Synthesis and Ribosome Structure. — Wiley-VCH, 2004. — С. 3. — ISBN 3-527-30638-2.
- А. С. Спирин. Биоорганическая химия. — М.: Высшая школа, 1986. — С. 10.
- Илья Леенсон. Язык химии. Этимология химических названий. — АСТ, 2016. — ISBN 978-5-17-095739-2.
- Jankowski JAZ, Polak JM. Clinical gene analysis and manipulation: tools, techniques and troubleshooting. — Cambridge University Press, 1996. — P. 14. — ISBN 0521478960.
- Barciszewski J, Frederic B, Clark C. RNA biochemistry and biotechnology. — Springer, 1999. — P. 73–87. — ISBN 0792358627.
- Spriggs KA, Stoneley M, Bushell M, Willis AE. (2008). «Re-programming of translation following cell stress allows IRES-mediated translation to predominate». Biol Cell. 100 (1): 27–38.
- ↑
- Wagner R., Theissen G., Zacharias. Regulation of Ribosomal RNA synthesis and Control of ribosome Formation in E.coli. — 1993. — P. 119–129.
- ↑ Cooper GC, Hausman RE. The Cell: A Molecular Approach. — 3rd edition. — Sinauer, 2004. — P. 261–76, 297, 339–44. — ISBN 0-87893-214-3.
- ↑
- ↑ Berg JM, Tymoczko JL, Stryer L. Biochemistry. — 5th edition. — WH Freeman and Company, 2002. — P. 118–19, 781–808. — ISBN 0-7167-4684-0.
- Gilbert SF. Developmental Biology. — 7th ed. — Sinauer, 2003. — P. 101–3. — ISBN 0878932585.
- ↑
- ↑
- ↑
- ↑
Функции
Первоначально Y-РНК были описаны как некодирующие РНК, связывающиеся с белком Ro60, который является антигеном, распознаваемым антителами из крови пациентов с системной красной волчанкой и синдромом Шегрена. Ro60 у позвоночных консервативен, его гомологи были выявлены у большинства Metazoa, а также в 5 % бактериальных геномов (среди которых геномы D. radiodurans и Salmonella). У всех этих организмов гомологи Ro60 связываются с Y-РНК или подобными им РНК с образованием рибонуклеопротеинов. У позвоночных Ro60 взаимодействует с нижним стеблевым доменом Y-РНК. С частью рибонуклеопротеинов, состоящих из Ro60 и Y-РНК (RoRNP), связывается белок La. Он необходим для правильной терминации транскрипции, опосредованной РНК-полимеразой III, и связывается с 3′-концевым хвостом новосинтезированных РНК в ядре. Большинство РНК впоследствии утрачивают полиуридиновый хвост, но Y-РНК его сохраняют и продолжают взаимодействовать с La. Он участвует в накоплении Y-РНК в ядре и защищает их от разрушения экзонуклеазами.
RoRNP участвуют в контроле качества некодирующих РНК, регулируют стабильность РНК и у некоторых видов участвуют в клеточном ответе на стресс. У многих видов, в числе которых лягушка Xenopus laevis, нематода C. elegans и домовая мышь, Ro60 связывается с такими дефектными некодирующими РНК, как неправильно уложенные 5S рРНК и . Нематоды, лишённые гомолога Ro60, жизнеспособны, однако у них нарушается процесс формирования особой личинки, предназначенной для перенесения неблагоприятных условий. Под действием УФ-излучения в клетках мыши и D. radiodurans происходит накопление RoRNP, и в отсутствие Ro60 их устойчивость к УФ-излучению понижается. У D. radiodurans ортолог Ro60 участвует в созревании рРНК, экспрессия которых была индуцирована тепловым стрессом, и разрушении рРНК, вызванном голоданием. Связывание Ro60 с неправильно уложенными некодирующими РНК неспецифично в отношении последовательности, и, по-видимому, этот белок может связываться с самыми разными РНК. По этой причине RoRNP можно рассматривать как внутриклеточные сенсоры стресса. В отсутствие Ro60 уровень Y-РНК в клетках как эукариот, так и прокариот снижался, из чего, вероятно, следует, что Ro60 стабилизирует Y-РНК, с которыми он взаимодействует.
Y-РНК участвует в инициации репликации хромосомной ДНК, а именно в формировании новых репликативных вилок. Однако они не нужны для элонгации и репликации и самого процесса удвоения ДНК как такового. Y-РНК вовлечены в инициацию репликации ДНК не сами по себе, а посредством белков. Для участия в инициации репликации ДНК необходим верхний стебель Y-РНК, но не участки связывания с белками Ro60 и La и не сами эти белки. Фактически около половины Y-РНК в человеческих клетках существует вне комплексов с белками Ro60 и La. Нарушение работы Y-РНК с помощью РНК-интерференции блокирует репликацию ДНК и пролиферацию клеток млекопитающих. При этом нормальный фенотип возвращается при введении в такие клетки короткой двуцепочечной РНК, соответствующей верхнему стеблю РНК hY1. Инактивация Y-РНК с помощью морфолиновых олигонуклеотидов в эмбрионах X. laevis и рыбы Danio rerio приводит к остановке развития и гибели на ранних этапах эмбриогенеза. В человеческих опухолях, напротив, наблюдается сверхэкспрессия Y-РНК по сравнению с нормальными тканями. Последовательности верхних стеблей Y-РНК таких организмов, как C. elegans, B. floridae и D. radiodurans, сильно отличаются от последовательностей позвоночных, и эти Y-РНК не могут функционально заменить Y-РНК в клетках человека.
Конкретный механизм участия Y-РНК в инициации репликации ДНК неизвестен. Известно, однако, что Y-РНК взаимодействует с несколькими белками, вовлечёнными в инициацию репликации ДНК, в числе которых репликации ORC, а также , и DUE-B, но не взаимодействует с белками репликативной вилки. С помощью Y-РНК удалось показать, что у человека Y-РНК динамически взаимодействуют с нереплицированным хроматином в G1-фазе клеточного цикла, колокализуясь с несколькими белками инициации репликации ДНК. После того как репликация началась, Y-РНК вытесняются из места начала репликации и не выявляются в местах активного синтеза ДНК.
Характеристики
Одноцепочечные РНК-вирусы и смысловая РНК
РНК-вирусы могут быть дополнительно классифицированы в соответствии с полярностью их РНК на и , или РНК . Вирусы, содержащие одноцепочечную (+)РНК похожую на мРНК и таким образом может непосредственно транслирована зараженной клеткой. Поэтому (+)РНК также называется смысловой. Антисмысловая (-)РНК вируса комплиментарна мРНК и таким образом может быть превращена в (+)РНК действием РНК-зависимой РНК-полимеразой перед трансляцией. Очищенная РНК (+)РНК-содержащих вирусов сама по себе может напрямую вызывать инфекцию, хотя ее инфекционность ниже, чем у полноценных вирусных частиц. Очищенная РНК (-)РНК-содержащих вирусов неинфекционна сама по себе, так как она должна транскрибироваться в (+)РНК; каждый вирион после транскрипции может быть источником нескольких смысловых цепей (+)РНК. Двухцепочечные(±)РНК вирусы напоминают (-)РНК-содержащие вирусы тем, что они также транслируют гены с отрицательной (некодирующей) цепи РНК.
Двуцепочечные РНК вирусы
Структура вириона реовируса
Двуцепочечные РНК вирусы представляет разнообразную группу вирусов отличающихся по заражаемому кругу хозяев (людей, животных, растений, грибов, и бактерий), числу фрагментов генома (от 1 до 12 сегментов), и организацией вириона (формой капсида, количеством слоев капсида, выросты, шипы, и т.п.). Члены этой группы включают ротавирусы, всемирно известные как наиболее распространенная причина детских гастроэнтеритов, и пикорнавирусы, известный во всем мире как наиболее часто встречающийся вирус в фекалиях людей и животных с признаками диареи или без них. является экономически важным возбудителем болезни коз и овец. В прошлые годы, достигнут большой прогресс в определении атомной и субнанометровой структуры ряда ключевых вирусных белков и капсидов вирионов нескольких дцРНК-вирусов, подчеркивая существенные параллели в структуре и механизмах процессов репликации многих из этих вирусов.
Частота мутаций
РНК вирусы имеют очень высокую частоту мутаций по-сравнению с ДНК-вирусами, потому что вирусная РНК-зависимая РНК-полимераза не имеет способности (3′-5′-экзонуклеазной гидролитической активности) к коррекции ошибок, как ДНК-полимеразы. Это одна из причин того, почему так трудно сделать эффективную вакцину для предотвращения болезней, вызываемых РНК-вирусами.
Ретровирусы также имеют высокую частоту мутаций, даже несмотря на то, что их промежуточная форма генома в виде ДНК интегрируется в геном хозяина (и, следовательно, после интеграции в геном, во время репликации, подвергается корректирующей проверке ДНК-полимеразой хозяина). Высокая частота мутаций обусловлена высокой частотой ошибок во время обратной транскрипции, до стадии интеграции в геном хозяина. Ошибки при этом встраиваются в обе цепи ДНК.
Некоторые гены РНК-вирусов важны для их репликации и мутации в них не допустимы. Например, участок генома вируса гепатита С кодирующий коровый белок, высоко консервативен, потому что он содержит структурную РНК, вовлеченную в образование участка внутренней посадки рибосомы.
Определение РНК гепатита С
Определение РНК вируса HCV-RNA, также именуемый ПЦР гепатита С, это исследование биологического материала (крови), с помощью которого можно определить в организме непосредственное наличие самого геноматериала вируса гепатита (любой отдельно взятый вирус является одной-единственной частичкой РНК).
Основной метод проведения теста — ПЦР, или метод полимеразно-цепной реакции.
Существуют два вида тестов крови на определение РНК HCV:
- качественный;
- количественный.
Качественный тест
Проведение качественного анализа дает возможность определить, находится ли вирус в составе крови. Все пациенты, у которых найдены антитела С-гепатита, должны пройти данный тест. По его результатам можно получить 2 ответа: «присутствует» либо «отсутствует» вирус. По положительному результату теста (обнаружено) можно судить об активном размножении вируса, который заражает здоровые клетки в печени.
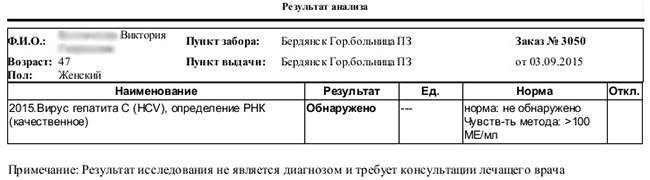
Тест, проводимый на качественный ПЦР, настроен на конкретную чувствительность, от 10 до 500 МЕ/мл. Если обнаруженный в составе крови вирус гепатита при удельном содержании меньше 10 МЕ/мл, то обнаружение вируса может стать невозможным. Очень низкое значение удельного содержания вируса наблюдается среди пациентов, для которых назначена противовирусная терапия. Поэтому важным является тот факт, насколько высока чувствительность медицинской системы для диагностирования и постановки качественного результата при полимеразной цепной реакции.
Зачастую полимеразная цепная реакция C-гепатита проводится сразу же после нахождения соответствующих антител. Последующие тесты, при прохождении противовирусной терапии, проводятся на 4-ю, 12-ю и 24-ю недели. И еще один анализ после прекращения ПВТ делается через 24 недели. Затем — один раз в год.
Количественный тест
Количественный анализ ПЦР РНК, иногда называемый вирусной нагрузкой, определяет концентрацию (удельное содержание) вируса в составе крови. Другими словами, под вирусной нагрузкой понимается определенное количество вирусной РНК, которое может находиться в конкретном количестве крови (принято использовать 1 мл, равный 1 см в кубе).

Единицы измерения для результатов теста — международные (стандартные) единицы, разделенные на один миллилитр (МЕ/мл). Содержание вируса иногда представляется по-разному, это зависит от лабораторий, где проводятся исследования. Для гепатита С количественное определение иногда использует такие значения, как копии/мл.
Проверка «вирусной нагруженности» позволяет определить степень инфекционности заболевания. Так риск заразить вирусом другого человека повышается при повышении концентрации гепатита в составе крови. К тому же высокое содержание вируса снижает эффект от проводимого лечения. Поэтому малая вирусная нагрузка является очень благоприятным фактором для успешного лечения.
К тому же тест гепатита С и его определение методом ПЦР играют большую роль при применении терапии от болезни и определения успешности лечения. На основании результатов теста осуществляется планирование курса реабилитации. Например, при слишком медлительном уменьшении удельной концентрации вируса гепатита, противовирусная терапия продлевается, и наоборот.

В современной медицине считается, что нагрузка больше 800000 ME/мл является высокой. Нагрузка же сверх 10000000 ME/мл считается критической. Но у специалистов из разных стран и по сей день не появилось одинакового мнения о пределах вирусной нагрузки.
Частота проведения количественного теста
В общих случаях количественный анализ на гепатит HCV-RNA делается перед проведением противовирусной терапии и через 3 месяца по окончании лечебных процедур для определения качества проведенной терапии.
В качестве результата для количественного теста станет считаться количественная оценка результатов по образцу, указанному выше. В результате будет вынесен вердикт «ниже измеряемого диапазона» или «в крови не обнаружено» — это является нормой для здорового человека.
Параметр чувствительности качественного теста обычно ниже, чем чувствительность количественного анализа. Расшифровка «Отсутствующий» показывает, что оба вида анализов не нашли РНК вируса. При показателе теста «ниже измеренного диапазона» анализ количественного типа, скорее всего, не нашел РНК гепатита, хотя это подтверждает наличие вируса с очень маленьким удельным содержанием.
Разнообразие РНК и их функций
РНК принято подразделять на кодирующие и некодирующие. Среди кодирующих РНК гл. место занимают мРНК. В соответствии с «центр. догмой» молекулярной биологии (ДНК – РНК – белок) они переносят генетич. информацию от ДНК к рибосомам, где она декодируется и реализуется в виде аминокислотных последовательностей белков. Кодирующей функцией обладает РНК-компонент фермента теломеразы (теломеразная РНК), определённый сегмент которой служит матрицей для синтеза теломерных ДНК-повторов на концах линейных хромосом. Со специфич. белками связана также короткая матричная РНК, называемая направляющей РНК (англ. guide RNA), участвующая в процессе редактирования мРНК. Она определяет, в какие участки мРНК будут введены дополнит. остатки U. У всех бактерий существует транспортно-матричная РНК (тмРНК), кодирующая короткий пептид, наращиваемый на С-концы дефектных белков, синтез которых по какой-то причине не смогли довести до конца рибосомы. Кодируемый тмРНК пептид служит сигналом для протеаз, уничтожающих дефектные белки. Т. о., в этом случае РНК осуществляет контроль качества белков, синтезируемых бактериальной клеткой. К кодирующим РНК относятся все вирусные РНК, которые служат матрицами для синтеза ДНК и РНК, а также вирусных белков.
К числу некодирующих РНК относится группа, формирующая белоксинтезирующий аппарат клетки. Более 80% РНК любой клетки представлено рибосомными рибонуклеиновыми кислотами (рРНК) – гл. структурными и функциональными компонентами рибосом.
Функцию декодирования генетич. информации на рибосомах под контролем рРНК осуществляют транспортные рибонуклеиновые кислоты (тРНК). Они переносят в рибосому аминокислотные остатки для синтеза белков. Когда рибосомы синтезируют секреторные или мембранные белки, с ними ассоциированы сигналузнающие РНК-белковые комплексы – т. н. SRP (от англ. signal recognition particles). В этих частицах РНК выполняют роль каркаса, на котором собираются белки, распознающие спец. сигнальные последовательности в синтезирующихся полипептидных цепях.
В регуляции процессов транскрипции и трансляции в клетке принимают участие многочисл. низкомолекулярные, или малые, и высокомолекулярные, или длинные, некодирующие РНК (мнкРНК и длнкРНК соответственно). Некоторые из них обладают рибозимной активностью и участвуют в процессинге др. РНК (напр., РНК-компонент бактериальной рибонуклеазы). В ядрах клеток эукариот присутствует семейство из 6–7 малых ядерных РНК (мяРНК), выполняющих гл. функцию на всех стадиях сплайсинга мРНК. Возможно, некоторые из них обладают рибозимной активностью. мнкРНК, локализующиеся в ядрышках эукариотич. клеток, отвечают за специфич. метилирование предшественников рРНК, за превращение в них определённых остатков уридина в псевдоуридин; они играют ключевую роль в РНК-интерференции. Многие из длнкРНК транскрибируются с цепи ДНК, противоположной кодирущей мРНК. Образуя комплементарные комплексы с мРНК вместе со специфич. белками, они полностью или частично подавляют экспрессию генов на уровне трансляции. Некодирующими РНК представлены транскрипты существенно большей части геномов всех организмов, чем кодирующими, т. е. разнообразие таких РНК в десятки раз превышает разнообразие индивидуальных белков в клетке.
Открытие у РНК столь высокой структурной и функциональной пластичности, в частности способности к самовоспроизведению и самопревращениям, позволяет предполагать, что при зарождении жизни на Земле эта нуклеиновая кислота была первым информац. биополимером, предшествовавшим появлению ДНК и белков.